

Рис. 1. Biolog EcoPlate — плашка для определения спектра пищевых ресурсов, используемых отдельным видом микроорганизмов или целым микробным сообществом. В каждой ячейке находится какое-нибудь вещество — потенциальный пищевой ресурс. По яркости окраски можно судить, с какой интенсивностью микробы, заселенные в плашку, используют данное вещество. С помощью таких устройств авторы обсуждаемой статьи оценивали пищевые спектры подопытных штаммов и функциональное разнообразие сообществ.
Эволюционные эксперименты на микробных сообществах, составленных из разного числа близкородственных видов, показали, что биоразнообразие может стимулировать свой собственный рост, способствуя диверсификации микробов. В основе обнаруженного механизма лежит конкуренция за пищевые ресурсы. По мере роста числа видов в сообществе обостряется конкуренция за узкий круг самых «популярных» источников углерода, тогда как другие источники остаются недоиспользованными. В результате получают селективное преимущество различные мутанты, у которых скорость размножения в отсутствие межвидовой конкуренции понижена, но зато повышена эффективность использования непопулярных субстратов. В сообществе с низким разнообразием такие мутанты отсеиваются отбором, и поэтому биоразнообразие простых сообществ так и остается низким, а сложных — растет.
В эволюции биоразнообразия задействованы как отрицательные, так и положительные обратные связи. В сообществе с высоким уровнем разнообразия дальнейшая диверсификация может быть заторможена из-за того, что все потенциальные ниши уже заполнены и все ресурсы поделены. Это пример отрицательной обратной связи. С другой стороны, рост разнообразия может ускорять сам себя благодаря таким явлениям, как создание ниш (Niche construction) и снижение вероятности вымирания видов, приуроченных к сообществам с высоким уровнем разнообразия. Это — примеры положительных обратных связей в эволюции биоразнообразия.
В серии краткосрочных эволюционных экспериментов на бактериях Pseudomonas fluorescens биологи из Германии и Франции продемонстрировали еще один потенциальный механизм, благодаря которому биоразнообразие может подстегивать свой собственный рост.
Эксперименты проводились на восьми штаммах псевдомонад, которые формально все относятся к виду Pseudomonas fluorescens, но при этом сильно различаются по своим биохимическим и экологическим параметрам, и поэтому авторы уверенно называют их «видами», игнорируя номенклатурную нестыковку.
В центре внимания исследователей был штамм (или вид) P. Fluorescens F113, в геном которого был добавлен ген устойчивости к канамицину, чтобы было удобно отделять его от остальных видов. Из этого вида и семи других составлялись микробные сообщества с разным уровнем видового и функционального разнообразия. Под функциональным разнообразием (ФР) понимается разнообразие спектров использования пищевых ресурсов (источников углерода) у бактерий, входящих в сообщество. Спектр питания каждого штамма определяли при помощи плашек Biolog EcoPlate (рис. 1).
Исследователи контролировали не только разнообразие сообщества, в котором шла эволюция штамма F113, но и способность бактерий к быстрым эволюционным изменениям. Для этого каждый эксперимент повторно проводился с модифицированными бактериями F113, у которых был отключен ген сайт-специфичной рекомбиназы XerD. Этот фермент время от времени перекраивает геном, меняя местами его фрагменты, что резко повышает частоту появления мутантных клеток. Поэтому его отключение должно снижать способность бактерий к быстрой адаптации.
В ходе каждого эксперимента свежеизготовленное микробное сообщество с тем или иным уровнем функционального разнообразия (ФР) и штамм F113 смешивались в пропорции 4:1, а полученная смесь затем эволюционировала в течение 48 часов, что соответствует примерно полусотне поколений (клеточных делений). Авторы надеялись, что за такое короткое время бактерии F113 rec− (не имеющие XerD), не успеют измениться, потому что у них будут слишком редко возникать полезные мутации. Эти ожидания впоследствии подтвердились.
Оказалось, что чем выше ФР сообщества, тем ниже доля F113 в итоговой смеси микробов по окончании 48-часового эксперимента. Это значит, что интенсивная межвидовая конкуренция за ресурсы ограничивает размножение F113. То же самое можно сказать другими словами: относительная приспособленность F113 (по отношению к остальному сообществу) снижается по мере роста ФР.
У микробов дикого типа (F113 rec+) снижение приспособленности с ростом ФР выражено слабее, чем у модифицированных микробов с замедленным мутагенезом (F113 rec−). Дополнительные опыты подтвердили, что микробы F113 rec+ по окончании эксперимента успешнее конкурируют с другими микробами из «своего» сообщества, чем F113 rec−. Это значит, что благодаря наличию рекомбиназы XerD адаптация бактерий к жизни в сообществе прошла успешнее.
Теперь нужно было проверить, происходила ли у F113 rec+ в ходе эксперимента диверсификация, то есть появлялись ли новые фенотипы, отличающиеся от исходного по спектру используемых ресурсов. Для этого взвесь бактерий разбавляли, высевали на агар и изучали морфологию колоний, развившихся из единичных клеток. Дело в том, что у P. fluorescens, как и у многих других бактерий, мутации, меняющие обмен веществ, нередко меняют также и форму колоний. Оказалось, что среди бактерий F113 rec+ действительно появилось много мутантов. При этом чем выше ФР сообщества, тем больше была доля различных мутантов среди F113 rec+ к концу 48-часового эксперимента. В ряде случаев мутанты полностью вытеснили исходный штамм (рис. 2). Таким образом, подтвердилось предположение о том, что разнообразие сообщества способствует диверсификации. Среди микробов F113 rec− мутантов обнаружить не удалось.
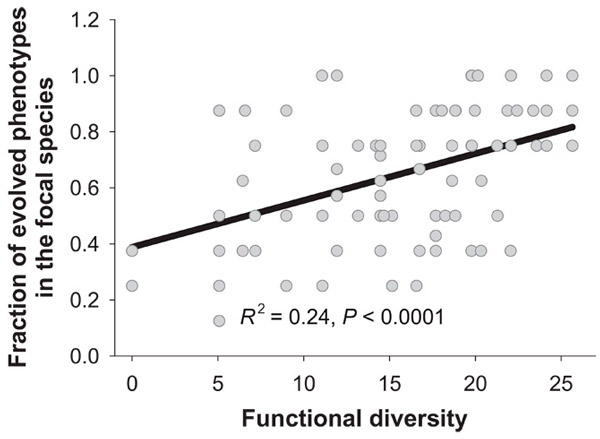
Рис. 2. Доля мутантных клеток среди бактерий F113 rec+ по окончании 48-часового эволюционного эксперимента в зависимости от функционального разнообразия сообщества, в котором шла эволюция.
Авторы определили спектр использования пищевых ресурсов бактериями-мутантами и сравнили его с пищевыми предпочтениями исходных бактерий F113, а также семи других видов, участвовавших в эксперименте. Оказалось, что основная конкуренция между микробами идет лишь за часть веществ, присутствующих в питательной среде. Этими «популярными» источниками углерода питается большинство видов. При этом в среде присутствует много других питательных веществ, которые используются менее интенсивно и лишь отдельными видами. Соответственно, конкуренция за эти «непопулярные» вещества слабее. Восемь видов, участвовавших в эксперименте, имеют похожие спектры использования «популярных» веществ, но сильно различаются по использованию «непопулярных».
Исходный штамм F113 был трофическим специалистом, то есть он, как и большинство других видов — участников эксперимента, в основном питался небольшим набором «популярных» веществ и лишь в минимальной степени использовал «непопулярные». Все бактерии-мутанты, произошедшие от F113 и достигшие заметной численности (то есть поддержанные отбором), отличаются от исходного штамма более широким спектром активно используемых ресурсов. Мутанты в значительной мере перешли на «непопулярные» источники углерода. По сравнению с предковым штаммом F113 они стали трофическими генералистами. Очевидно, это помогло им избежать конкуренции и обеспечило успех в сообществах с высоким ФР.
Выяснилось также, что мутанты, научившиеся эффективно утилизировать недоиспользуемые сообществом ресурсы, заплатили за это эволюционное достижение немалую цену: их приспособленность к жизни в условиях пониженной межвидовой конкуренции снизилась. Они проигрывают в скорости роста своим предкам — немутантным бактериям F113, если поблизости нет других видов-конкурентов (или если таких видов мало и ФР сообщества низкое). Это объясняет, почему мутанты — пищевые генералисты распространились в основном в сообществах с высоким ФР, где приспособленность исходного штамма F113 оказалась понижена из-за острой межвидовой конкуренции. Только поэтому мутанты-генералисты и смогли с ним тягаться.
В том, что за успешную утилизацию одних веществ пришлось заплатить снижением эффективности использования других, ничего неожиданного нет: это общее, хотя и имеющее множество исключений, эволюционное правило — принцип «эволюционного компромисса». Специалист, как известно, подобен флюсу, но в своей узкой области он, как правило, побеждает в конкуренции генералиста. Важно, что в данном случае удалось экспериментально показать роль эволюционных компромиссов в формировании разнообразных сообществ. Если бы штаммы, способные эффективно утилизировать «непопулярные» источники углерода, не проигрывали исходному штамму в конкуренции за «популярные» субстраты или за что-то другое в иных условиях, они бы давно вытеснили его повсеместно, и глобальное биоразнообразие снизилось бы.
Таким образом, исследование подтвердило, что высокое разнообразие сообщества может стимулировать свой собственный рост, способствуя диверсификации входящих в сообщество видов. Предложенный механизм основан на конкуренции за ресурсы и отличается от «создания ниш», хотя авторы и признают, что создание ниш (например, использование одними штаммами продуктов жизнедеятельности других или иные варианты взаимовыгодных отношений), тоже могло внести какой-то вклад в полученные результаты.
Очевидно, что рост биоразнообразия не может ускорять диверсификацию до бесконечности: в какой-то момент должно наступить насыщение. Вся игра строится на наличии в системе недоиспользуемых ресурсов, а рано или поздно биоразнообразие вырастет настолько, что ресурсы будут поделены до последней крошки. Скорее всего, биоразнообразие начнет тормозить диверсификацию задолго до этого момента. Дальнейшие исследования покажут, насколько широко распространен в природе данный механизм автокаталитического роста биоразнообразия.
Помимо прочего, исследование наглядно показало, что «экологический» и «эволюционный» масштабы времени практически совпадают, по крайней мере в мире микробов. Самые настоящие эволюционные изменения, основанные на мутациях и отборе, меняющие функциональную структуру и разнообразие сообщества, происходили в этом эксперименте за какие-то смешные 48 часов.
Источник: ЭЛЕМЕНТЫ БОЛЬШОЙ НАУКИ
