
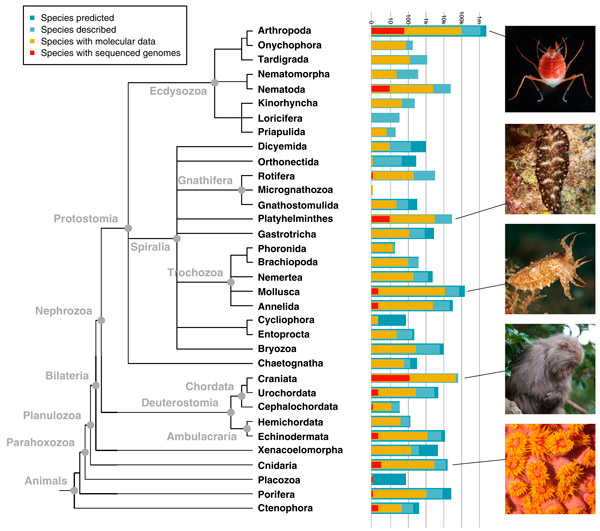
Рис. 1. Генетические данные на фоне современного эволюционного древа животных. Столбики против названий типов показывают число видов: синий — предсказанное, голубой — реально описанное, желтый — виды, по которым есть хоть какие-то молекулярно-генетические данные, красный — виды, у которых прочитаны полные геномы.
Сравнение полных геномов — важнейший метод изучения эволюции, ставший доступным биологам с 1995 года. К настоящему времени полностью прочитано 212 геномов многоклеточных животных. Это уже позволило сделать ряд общих выводов — например, частично реконструировать минимальный геном последнего общего предка животных, а также оценить характерную для современных геномов долю новых генов, свойственных только отдельным систематическим группам.
«Сравнение полных геномов представляет собой единственный удовлетворяющий исследователя путь к реконструкции эволюции», — пишет в книге «Логика случая» знаменитый биоинформатик Евгений Кунин. С категоричностью этого утверждения можно не согласиться, но речь идет и правда о важнейшей вещи. Сравнение полных геномов — метод, который дал биологам совершенно новые возможности для наблюдения эволюционного процесса.
Эпоха сравнения полных геномов началась в 1995 году, когда лаборатория Крейга Вентера опубликовала результат прочтения генома бактерии гемофильного гриппа Haemophilus influenzae. В течение года с того момента было прочитано еще несколько геномов бактерий и один эукариотный — геном дрожжей Saccharomyces cerevisiae. Их можно и нужно было сравнивать, разрабатывая для этого приемы, фиксируя открывающиеся закономерности. За считанные годы родилась новая наука — сравнительная геномика, подобно тому, как в XVIII веке сложилась сравнительная анатомия.
Несколько недель назад американские биологи Кейси Данн и Джозеф Райан выпустили очень краткий — на пяти страничках — обзор данных, полученных из прочитанных на данный момент полных геномов многоклеточных животных. Их уже столько, что можно сделать первые выводы, касающиеся генетической эволюции животных в целом.
Данн и Райан определили предмет своего интереса совершенно четко: макроэволюция ядерных геномов. Оговорка о макроэволюции означает, что нас сейчас интересуют геномные различия между крупными группами организмов, а не те, что можно найти между близкими видами, и тем более не внутривидовые. Оговорка про ядерные геномы означает, что мы не рассматриваем, например, митохондриальную ДНК (которую тоже очень интересно изучать, но это особая тема). Итак, что нам известно о ядерных геномах животных?
Сначала сухие цифры. На момент публикации статьи Данна и Райана число полностью прочитанных геномов животных составляло 212 (даже за прошедшее с тех пор короткое время их могло стать уже больше). Подразумевается, что каждый геном принадлежит отдельному виду. Двести двенадцать видов — это меньше двух сотых долей процента от общего числа видов современных животных, которое составляет примерно 1,5 миллиона. Кроме того, разные группы животных представлены в этой выборке очень неравномерно. Во-первых, 83% прочитанных геномов принадлежат позвоночным. Во-вторых, наблюдается явный перекос в сторону животных с маленькими геномами, работать с которыми технически гораздо проще. Напомним, что размеры геномов разных животных могут отличаться на три порядка, то есть в тысячи раз. Обладателям гигантских геномов, например, двоякодышащим рыбам, придется еще какое-то время подождать своей очереди.
Тем не менее, все крупные эволюционные ветви животных современной выборкой охвачены, так что ее можно считать вполне представительной (рис. 1).
Уточним это утверждение. На эволюционном древе есть семь крупных ветвей, без упоминания которых не обходится ни один разговор о животном царстве (если, конечно, его хотят охватить в целом). Это губки, гребневики, пластинчатые, стрекающие, линяющие, спиральнодробящиеся и вторичноротые. Так вот, во всех этих ветвях без исключения есть представители, полные геномы которых уже прочитаны и внесены в электронные базы. В большинстве ветвей — и не по одному.
Все геномы животных гораздо более похожи друг на друга, чем можно было бы ожидать, исходя из внешних различий их обладателей. Например, размер генома очень слабо коррелирует со сложностью организма. Конечно, упоминая в биологии «сложность», приходится всегда оговаривать, что понятие это абстрактное и трудно определимое. Но при любом его понимании не приходится сомневаться, например, в том, что человек, одна только нервная система которого насчитывает 86 миллиардов нейронов, сложнее круглого червя Caenorhabditis elegans, у которого нейронов 302 штуки. В догеномную эпоху высказывались предположения, что у человека примерно 140 тысяч генов, но сейчас ясно, что их всего 20–25 тысяч. А число генов ценорабдитиса — 19 735. По числу генов человек и ценорабдитис мало отличаются друг от друга.
Любой геном можно охарактеризовать двумя главными параметрами: размер генома (C) и число генов (G). Размер генома измеряется в парах нуклеотидов, число генов — просто в штуках. Широко известно, что размер генома может очень сильно — на порядки — отличаться у организмов, явно близких по сложности устройства тела (например, у хвостатых и бесхвостых земноводных). Это явление называют «парадоксом значений C».
А вот с числом генов все наоборот. Оно часто бывает примерно одинаковым (во всяком случае, сравнимым) у организмов, отличающихся чуть ли не по всем признакам устройства тела, какие только можно придумать. Этот факт получил название «парадокса значений G». Теоретически объяснять его можно по-разному, но в любом случае это один из главных выводов, сделанных с помощью чтения полных геномов. Другими способами надежно узнать число генов нельзя. Например, если мы измеряем химическими методами количество ДНК в клетке, то сразу получаем информацию о размере генома, но отличить гены от некодирующих последовательностей (которых в геномах эукариот всегда много) таким образом невозможно.
Еще одна задача, которую нельзя решить без исследований полных геномов — это реконструкция генома общего предка всех многоклеточных животных. Сравнение геномов разных животных, а также их одноклеточных родственников, позволяет в принципе определить минимальный набор генов, которыми этот предок должен был обладать. На данный момент можно сказать, что у последнего общего предка всех животных было по меньшей мере 6289 генных семейств, общих у него с некоторыми одноклеточными, и 2141 генное семейство, уникальное для животных (напомним, что генное семейство — это группа генов, происходящих от одного гена-предшественника путем его удвоений). Получается, что у общего предка животных было минимум 8430 генов. Правда, это — в случае, если каждое генное семейство состояло только из одного гена, что маловероятно. На самом деле генов, скорее всего, было больше, но все же эти цифры дают какую-никакую точку отсчета. И в любом случае, они не окончательные — ведь анализ геномов продолжается.
Некоторые выводы касаются отдельных групп генов и белков. Например, белки клеточной адгезии (cell adhesion molecules, CAM), обеспечивающие сцепление и взаимодействие клеток друг с другом, оказались вовсе не уникальными для животных — они появились у наших одноклеточных родственников, которым были нужны, вероятно, ввиду их сложного жизненного цикла. А вот многие гены и белки, обеспечивающие внутриклеточную сигнализацию, для животных как раз уникальны.
От генома последнего общего предка многоклеточных животных, естественно, произошли — путем многократных копирований с изменениями — геномы всех животных, доживших до современности. Так и сформировалось ветвистое эволюционное древо, которые мы теперь видим. События, меняющие геномы в ходе эволюции, очень разнообразны: дупликации разных типов, перестройки устойчивого совместного расположения генов (синтении), умножение или потеря генов тех или иных семейств, всевозможные вставки, модификации регуляторных областей ДНК, перенос мобильных генетических элементов, изменение состава повторяющихся последовательностей, таких как сателлитная ДНК, и многое другое. Все эти события оставляют следы, которые наслаиваются друг на друга. Их можно найти и прочитать, примерно так же, как можно найти и прочитать следы правок в обычном тексте. Ведь геном — тоже текст (хотя и не только текст, конечно). Геном любого животного заключает в себе целую летопись изменений, многие из которых можно довольно точно распознать и датировать. Сплошь и рядом это относится даже к изменениям, которые произошли много сотен миллионов лет назад. Текст может сохранить все.
Сейчас широко известно, что генный «инвентарь» многоклеточных животных эволюционно очень консервативен. Сплошь и рядом одни и те же гены используются для близких функций и у человека, и у относительно простых существ наподобие того же ценорабдитиса. Однако это — далеко не вся правда. В любом эукариотном геноме есть 10–20% генов, не похожих ни на какие гены смежных (и любых других) групп организмов. Такие гены часто называют «генами-сиротами» (orphan genes), имея в виду, что для них не удается найти генов-предков. Но все-таки корректнее называть их таксономически ограниченными генами (taxonomically-restricted genes, TRG). Таксономически ограниченный ген — это действительно новый ген, появившийся в эволюции одной-единственной группы организмов. Например, есть гены, свойственные только позвоночным, только некоторым насекомым или даже только мышам. Естественно, тут возникают интересные вопросы. Во-первых, откуда новые гены берутся? И, во-вторых, какие у них обычно бывают функции?
На вопрос, откуда берутся новые гены, самый распространенный ответ простой: новые гены берутся из старых. Например, в результате случайной дупликации образуются две копии некоего гена, в первой из них мутации накапливаются медленнее, во второй быстрее, в итоге вторая копия постепенно становится новым геном с новой функцией. Это действительно очень частый механизм создания новых генов, но не единственный. По-видимому, новые гены в принципе могут образоваться из чисто регуляторных участков ДНК или даже из некодирующих последовательностей. Иногда на это накладываются дополнительные события, например, вставка в новый ген небольшого куска другого, старого гена. Кстати, установлено, что «молодые» гены обычно кодируют более короткие белки, чем «старые». Получается, что долгоживущий ген как бы накапливает груз, состоящий из случайных вставок — которые, впрочем, в свою очередь могут подвергаться отбору и обретать новые функции. В любом случае, образование новых генов, как и большинство процессов в биологии, не сводится к какому-либо одному механизму. Таких механизмов несколько, и они могут сочетаться.
Что касается функций таксономически ограниченных генов, то они бывают очень разнообразными. Роднит их одно: большинство этих функций — специфические для данной группы животных, не встречающиеся за ее пределами.
Например, свой набор таксономически ограниченных генов есть у типа стрекающих, к которому относятся медузы и полипы (включая гидр и кораллов). Пожалуй, самая яркая особенность стрекающих животных — стрекательные клетки (книдоциты), в честь которых этот тип получил свое название. Книдоциты — одни из самых сложных клеток, какие только можно найти в животном мире. В каждом книдоците находится свернутая стрекательная нить с ядовитым веществом и механизмом выбрасывания. Книдоциты стрекающих очень разнообразны, их насчитывается более 30 типов, а вот ни у каких других животных подобных клеток нет. Так вот, оказалось, что очень многие гены, свойственные только стрекающим, связаны со специфическими органеллами стрекательных клеток. Например, целая группа таких генов кодирует миниколлагены — уникальные короткие белки, входящие в состав стенки внутриклеточной капсулы, в которой размещается стрекательная нить. У одного из видов пресноводной гидры найден 41 ген, активный только в стрекательных клетках и не похожий ни на какие гены других типов животных. Выходит, в общем логично: новые гены обеспечивают эволюционные новшества, проявляющиеся уже на уровне организма (рис. 2).
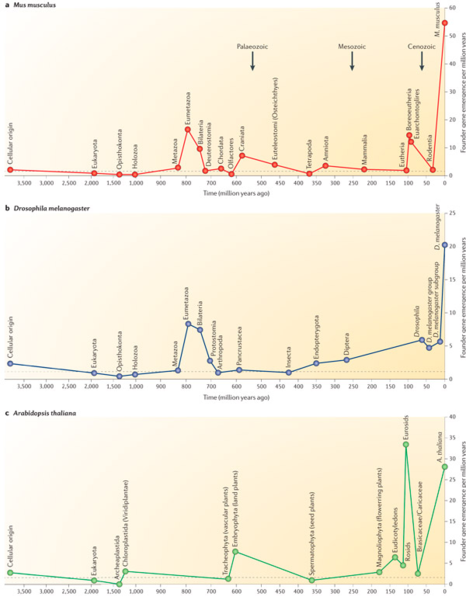
Рис. 2. Скорость появления новых генов в геномах предков домовой мыши (Mus musculus), мухи-дрозофилы (Drosophila melanogaster) и цветкового растения Arabidopsis thaliana. На каждом графике по горизонтали время (в миллионах лет назад), а по вертикали число новых генов, появившихся в среднем за миллион лет. Новые гены здесь называют также генами-основателями (founder genes), имея в виду, что они дают начало целым генным семействам. На графиках особенно интересны пики. В истории геномов животных хорошо заметен пик, связанный со становлением многоклеточности (Eumetazoa). В истории генома мыши есть еще два явных пика — в предполагаемые моменты возникновения позвоночных или черепных (Craniata) и плацентарных млекопитающих (Eutheria). В эти относительно краткие периоды появилось много новых генов. Свои пики есть и в истории генома зеленого растения. Вот чтобы составлять такие графики, знание полных геномов и нужно.
Любой геном — это в некотором смысле архив, документирующий события, происходившие на всем протяжении эволюционной линии от последнего общего предка всех живых организмов (last universal common ancestor, LUCA) до обладателя этого генома. Например, в геноме мыши запечатлены следы событий, происходивших по всей непрерывной цепочке предков и потомков от LUCA до этой самой мыши. Конечно, некоторые следы в ходе эволюции стираются бесследно, и с этим ничего не поделаешь. Но очень многие сохраняются. Причем все эти следы заключены внутри сложной упорядоченной структуры, дающей возможность их послойно датировать. В этом отношении работа сравнительного геномика напоминает стратиграфию или археологию.
Источник: ЭЛЕМЕНТЫ БОЛЬШОЙ НАУКИ
