

Рис. 1. Атлантический мечехвост Limulus polyphemus.
Полногеномная дупликация (whole genome duplication, WGD) долгое время считалась уникальной особенностью позвоночных животных, в ранней эволюции которых она, по-видимому, произошла дважды. Однако последние исследования показывают, что очень похожее неоднократное удвоение генома случилось и в одной эволюционной ветви членистоногих, а именно у мечехвостов. Появление в результате серии дупликаций большого числа новых генов может рассматриваться как одна из вероятных причин эволюционного успеха позвоночных, но мечехвостов оно к успеху не привело.
Знакомство с генами
Около полувека назад работавший в США японский биолог Сусуму Оно всерьез задумался над вопросом: откуда берутся новые гены?
Понятие «ген» к тому времени было знакомо ученым давно — Вильгельм Людвиг Иогансен ввел его еще в 1909 году. С тех пор все знали, что ген — это некая «единица наследственности». Но в первые десятилетия у биологов не было никакого четкого представления о том, что представляют собой гены физически. Очень долго никто даже не знал точно, из какого вещества они состоят (роль ДНК в передаче генетический информации удалось четко продемонстрировать только в 1944 году). В этих условиях многие гипотезы об эволюции генов оставались труднопроверяемыми. Например, Эрнст Майр — крупнейший эволюционист, знаменитый исследователь видообразования — твердо считал, что у разных организмов, даже близкородственных, сами наборы генов должны очень сильно отличаться. Но так ли это в действительности? И как образуются новые гены, если уж они настолько изменчивы?
В 1953 году Джеймс Уотсон и Фрэнсис Крик совершили самое знаменитое открытие в биологии XX века, установив химическую природу гена. Стало окончательно ясно, что ген — это участок двуспиральной молекулы ДНК, несущий (как правило) информацию о структуре одного определенного белка. ДНК представляет собой цепочку нуклеотидов, белок — цепочку аминокислот, при этом последовательности нуклеотидов в гене и аминокислот в белке связаны через генетический код. Между тем «читать» аминокислотные последовательности белков биохимики к 1950-м годам уже умели. Это стало хотя и косвенным, но очень надежным способом получения информации о структуре генов, способным во многих задачах заменить прямое чтение последовательностей ДНК (которое было освоено несколько позже). Родилась сравнительная молекулярная биология.
Первые же исследования такого рода показали, что гены в эволюционном масштабе времени поразительно консервативны. Например, в начале 1960-х годов биохимик Эмануэль Марголиаш изучил аминокислотную последовательность цитохрома c — широко распространенного белка, выполняющего важную функцию в процессе клеточного дыхания. Оказалось, что у всех изученных позвоночных этот белок состоит из 104 аминокислот, из которых, например, у человека и лошади различаются только 12, а у человека и тунца — только 21. Даже у пекарских дрожжей, которые вообще относятся не к животным, а к грибам, около половины аминокислотной последовательности цитохрома c совпало с такими же последовательностями, прочитанными у позвоночных. Другие подобные исследования дали близкие результаты: последовательности генов, кодирующих «один и тот же белок» (то есть белки с одинаковым действием и сходными свойствами), в разных организмах оказались очень похожими.
Таким образом, предсказание Эрнста Майра о большой генетической разнице между родственными организмами было опровергнуто прямыми наблюдениями. По словам известного биоинформатика Евгения Кунина, «ошибочность этого предсказания оказалась просто феерической, что само по себе делает его нетривиальным и ценным». На самом деле гены сплошь и рядом гораздо более устойчивы, чем организмы, в состав которых они входят.
Столь высокая устойчивость генов, на первый взгляд, делает достаточно маловероятным такое явление, как постепенная эволюция уже существующего гена, приводящая к полной перемене его функции. Но тогда откуда новые гены вообще берутся? Именно этим вопросом Сусуму Оно и задался.
Дупликации и эволюция
Японский генетик рассуждал следующим образом. Мы знаем, что ген, «нагруженный» важной функцией, от которой нельзя отказаться, может меняться в ходе эволюции лишь ограниченно, а главное, медленно. Если бы все гены эволюционировали с такой скоростью, как гены цитохрома у позвоночных, наблюдаемое нами сейчас разнообразие жизни было бы недостижимо. А поскольку оно все-таки есть, значит, должен существовать какой-то механизм ускорения эволюции на генном уровне.
Сусуму Оно предположил, что таким механизмом служат генные дупликации, то есть попросту удвоения. Возникающие в результате этого процесса дополнительные копии генов накапливают различия гораздо легче и свободнее, чем уникальные гены, — ведь их «порча» перестает быть опасной для жизни. И вот такая избыточная копия вполне может проэволюционировать до состояния нового гена за разумное время. Новизна создается через избыточность.
Свидетельства реальности такого способа эволюции были в распоряжении биологов уже в 1960-е годы. Например, белок, переносящий кислород в нашей крови, — гемоглобин — это огромная молекула, состоящая из четырех аминокислотных цепочек, которые синтезируются отдельно. В состав гемоглобина взрослого человека обычно входят аминокислотные цепи двух типов, а именно две α-цепи и две β-цепи. Известно, что α-цепь и β-цепь гемоглобина кодируются разными генами, причем расположенными в геноме далеко друг от друга (в разных хромосомах). Это явный результат дупликации единого гена, случившейся у далекого предка — около 500 миллионов лет назад, если судить по числу аминокислотных замен.
Надо заметить, что в истории с гемоглобинами Оно даже недооценил размах событий. Сейчас мы знаем, что на самом деле в геноме человека находится тринадцать генов гемоглобина, кодирующих аминокислотные цепи девяти разных типов. Многие из этих генов или активны только до рождения, или дают очень низкие концентрации продуктов, но, тем не менее, они существуют. Так что дупликация генов гемоглобина происходила в нашей эволюционной линии не единожды и не дважды.
Впрочем, Оно не ограничился изучением гемоглобинов. Он довольно быстро собрал другие подобные данные, касавшиеся, например, некоторых ферментов обмена веществ и белков иммунитета (иммуноглобулинов). В том, что генные дупликации служат важной движущей силой эволюции, он был убежден. Оставалось понять, как именно они происходят.
По одному или сразу?
Классическая генетика знает два относительно безопасных способа образования дополнительных копий генов: удвоение отдельного участка хромосомы (тандемная дупликация) или удвоение всего генома целиком. Оно прекрасно знал, что оба эти механизма вполне реальны, и был уверен, что оба они участвуют в крупномасштабной эволюции. Вопрос был лишь в том, какой из них важнее.
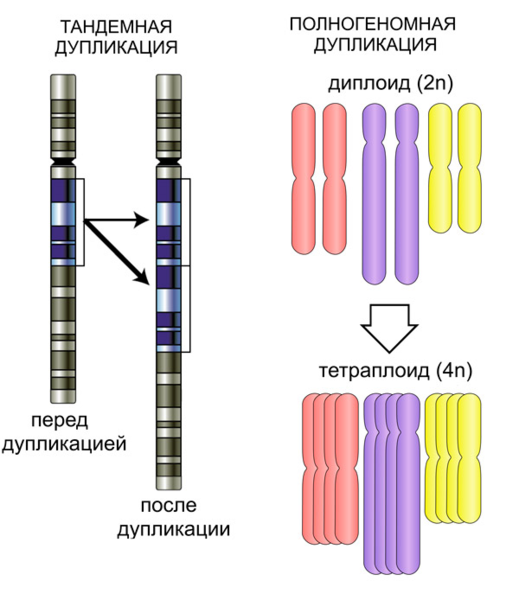
Рис. 2. Тандемная и полногеномная дупликации. Слева показана одна хромосома с дифференциальной окраской разных участков (один участок удваивается), справа — набор хромосом, который удваивается целиком.
Оценивая вероятности тех или иных событий, эволюционные биологи обычно исходят из закономерности, которую можно сформулировать просто: структура «боится» новизны. Чем событие «катастрофичнее» (в каком угодно смысле), тем выше риск того, что оно так или иначе нарушит функционирование сложной системы, и тем менее вероятно, что подобные события будут происходить в эволюции часто. Разница между полногеномной и тандемной дупликацией с этой точки зрения очевидна: первая затрагивает весь геном, вторая — только одну из хромосом, и то не целиком. Естественно, традиционно мыслящие биологи считали полногеномную дупликацию гораздо более редким механизмом (тем более что у животных, в отличие от растений, она и правда редко встречается).
Однако не всё так просто. После тандемной дупликации гены, которые в результате нее приобрели дополнительные копии, начинают синтезировать больше продуктов, в то время как остальные гены работают по-прежнему. В результате эволюционно выработанное соотношение продуктов разных генов нарушается, и отлаженная генетическая система может давать сбои. Между тем при полногеномной дупликации такие сбои исключены, потому что в этом случае число копий всех генов увеличивается пропорционально. И отсюда Оно сделал вывод: при прочих равных условиях полногеномная дупликация является более безопасным способом эволюции, чем множество тандемных дупликаций, дающих сравнимый с ней суммарный эффект.
Заметим, что этот суммарный эффект должен состоять не более и не менее как в увеличении всего генома минимум вдвое. Не пустой ли это разговор? Есть ли основания полагать, что такие события вообще происходили в эволюции каких бы то ни было животных?
Да, есть. Чтобы понять это, достаточно сравнить размеры геномов позвоночных и их ближайших родственников — низших хордовых.
Как раз на эту тему к 1960-м годам было собрано уже достаточно много данных. И данные эти однозначно указывали на следующее: при переходе от низших хордовых к позвоночным размер генома увеличился скачком. Это легко выразить в цифрах. У типичного позвоночного (на роль которого можно с одинаковым успехом выбрать акулу, лягушку, крысу или человека) геном почти наверняка окажется в 3–5 раз больше, чем у образцового низшего хордового — ланцетника. Есть отдельные группы позвоночных, у которых геном увеличен дополнительно (например, хвостатые амфибии) или, наоборот, уменьшен (например, птицы), но всё это выглядит как вариации на фоне уже достигнутого общего уровня.
Обдумывая эти данные, Оно пришел к очень смелой по тем временам гипотезе: в начале эволюции позвоночных у них произошло удвоение всего генома. А судя по тому, во сколько раз геном увеличился, таких событий могло быть и два.
Возможно ли такое чисто биологически? Да. Наука отлично знает (и знала уже в 1960-е годы) примеры позвоночных, у которых удвоение всего генома действительно произошло.
Еще в первые годы XX века было установлено, что у подавляющего большинства многоклеточных животных клетки (кроме половых) диплоидные, то есть содержат по две копии каждой хромосомы. Иногда наборы генов могут удваиваться. Тогда клетки, и состоящие из них организмы, становятся тетраплоидными — содержащими по четыре копии каждой хромосомы. Тетраплоидные расы есть, например, у серебряного карася — вида, к которому относится аквариумная золотая рыбка.
Бывает и так, что набор генов удваивается дважды подряд. Тогда организм становятся октоплоидным — содержащим по восемь копий каждой хромосомы. Октоплоидные виды с очень большим количеством хромосом есть, например, среди южноамериканских рогатых лягушек рода Ceratophrys.
 Рис. 3. Некоторые обладатели полногеномной дупликации: коловратка Adineta vaga (слева) и украшенная рогатка Ceratophrys ornata. У первой геном увеличен вдвое (по сравнению с типичным геномом коловраток), у второй — вчетверо (по сравнению с типичным геномом лягушек).
Рис. 3. Некоторые обладатели полногеномной дупликации: коловратка Adineta vaga (слева) и украшенная рогатка Ceratophrys ornata. У первой геном увеличен вдвое (по сравнению с типичным геномом коловраток), у второй — вчетверо (по сравнению с типичным геномом лягушек).
Если такое возможно, то почему бы и общему предку позвоночных не быть тетраплоидом или даже октоплоидом? Все следующие тридцать лет своей жизни Сусуму Оно хранил верность этой гипотезе, постоянно собирая новые данные в ее поддержку. Но сторонников у него было немного.
Дупликация и уникальность
В 1990-х годах британский генетик Питер Холланд заинтересовался эволюцией довольно специфической группы генов многоклеточных животных, которая называется Hox-генами. Эти гены активно участвуют в эмбриональном развитии, контролируя формирование различий между отделами тела. Одна из особенностей Hox-генов состоит в том, что они, как правило, располагаются в хромосомах компактными группами — кластерами. Так вот, в процессе исследований быстро выяснилось, что большинство групп позвоночных, например млекопитающие, имеет четыре Hox-кластера. А вот у животных, не относящихся к позвоночным, в том числе и у низших хордовых, Hox-кластер, как правило, только один. Почему?
В поисках ответа на этот вопрос Холланд решил расширить круг рассматриваемых генов — и сразу обнаружил ряд интересных вещей.
Во-первых, к началу 2000-х годов генетика позволяла уверенно утверждать, что у позвоночных больше генов, чем у каких угодно беспозвоночных, не исключая и низших хордовых. Подчеркнем: тут речь идет уже не о размере генома (который может зависеть и от объема «бессмысленных» некодирующих последовательностей), а именно о числе генов. Правда, общее число генов в геноме не всегда легко точно оценить, но как раз тут данные по отдельным группам генов бывают достаточно показательны: например, у ланцетника 15 Hox-генов, а у человека 39.
Во-вторых, оказалось, что не только Hox-гены, но и многие другие гены (сплошь и рядом никак не связанные с регуляцией эмбрионального развития) находятся в геномах ланцетника и других низших хордовых в единственном экземпляре, а в геномах позвоночных — в двух, трех или четырех слегка различающихся копиях. Список таких генов оказался очень длинным, в него вошли гены многих ферментов, сигнальных факторов, белков цитоскелета.
В 2002 году Питер Холланд в соавторстве с Ребеккой Фёрлонг опубликовал статью под недвусмысленным названием: «Были ли позвоночные октоплоидными?». И ответ был — да, были. Гипотеза о неоднократном удвоении всего генома позвоночных получила признание.
Надо отдать должное Холланду: в своих статьях он первым делом подробно разобрал аргументы Оно, похвалив его книгу (действительно очень интересную) и показав, в чем он был прав. В свою очередь, сам Оно никогда не оставлял без внимания новые генетические данные, в том числе и касавшиеся Hox-генов (в его последних статьях есть ссылки на того же Холланда). К сожалению, Сусуму Оно умер в начале 2000 года, совсем чуть-чуть не дожив до времени, когда его гипотеза стала полноценным научным мейнстримом.
Сейчас общепризнано, что в начале эволюции позвоночных у них произошла полногеномная дупликация. Причем на самом деле полногеномных дупликаций было две: вероятнее всего, одна из них случилась при самом возникновении позвоночных, а вторая — при переходе от бесчелюстных к челюстноротым. Так что октоплоидами можно считать только последних (рыб, амфибий, рептилий, млекопитающих).
Нужно заметить, что ни у каких позвоночных количество генов не превосходит таковое ланцетника (или других низших хордовых) ровно в четыре раза. Это превосходство всегда гораздо меньше. Дело в том, что две, а тем более четыре совершенно одинаковые копии функционального гена не только избыточны для организма, но и могут мешать друг другу работать. В лучшем случае эти копии начинают быстро накапливать различия и разделять между собой функции. В худшем же случае «лишние» гены выходят из строя, сначала превращаясь в псевдогены, а потом вообще исчезая. Так происходит предсказанная еще Оно вторичная диплоидизация: полиплоидный геном уменьшается, компактизуется и в итоге вновь становится диплоидным. Этот процесс очень хорошо изучен у растений, но и у животных он тоже, без сомнения, происходил.
Неудивительно, что в результате работ Холланда полногеномная дупликация стала считаться уникальным признаком группы позвоночных, таким же четким, как головной мозг, состоящий из пяти отделов, нервный гребень и некоторые другие анатомические структуры. Казалось ясным, что позвоночные — единственная крупная эволюционная ветвь животных, в которой произошло такое событие.
Тайна генома мечехвоста
Не вызывает сомнений, что полногеномные дупликации, охватывающие крупные эволюционные ветви, происходили в истории многоклеточных животных крайне редко. Кроме позвоночных нечто подобное до сих пор было обнаружено только у бделлоидных коловраток — очень своеобразных существ, полностью отказавшихся от полового размножения.
Однако в биологии никогда не следует спешить с обобщениями. В последние годы появились данные о наличии дополнительных копий ряда генов, родственных Hox-генам (но не обязательно к ним относящихся), у некоторых пауков, скорпионов и мечехвостов. Все эти животные относятся к древней эволюционной ветви, которая называется хелицеровыми. Не было ли в истории хелицеровых полногеномных дупликаций, подобных тем, что произошли у позвоночных?
Действительно, в прошлом году вышло подробное исследование генома атлантического мечехвоста, вполне подтверждающее эту идею. В частности, оказалось, что у мечехвоста не один Hox-кластер, как у подавляющего большинства беспозвоночных, а четыре, расположенных в разных хромосомах, — как у млекопитающих!
Неудивительно, что этим вопросом тут же занялась международная группа генетиков, в состав которой вместе с несколькими китайскими учеными вошел сам Питер Холланд. Прежде всего им надо было выяснить, является ли полногеномная дупликация уникальным признаком вида, к которому относится атлантический мечехвост, или же это особенность эволюционного ствола мечехвостов в целом. Для этого в исследование было включено еще два вида: мангровый мечехвост и китайский мечехвост. Вывод оказался однозначным: полногеномная дупликация — особенность всех современных мечехвостов, скорее всего унаследованная ими от общего предка.

Рис. 4. Мангровый мечехвост Carcinoscorpius rotundicauda (a), эволюционное древо современных мечехвостов (b) и некоторые гены трех видов мечехвостов (c). MYA — миллионы лет назад. В таблице показаны гены четырех групп; каждый столбик соответствует определенному гену, каждая строчка — одному из трех исследованных видов мечехвостов. Числа в клеточках показывают, сколько копий данного гена есть в геноме данного вида. Жирные цифры соответствуют генам, учтенным при построении молекулярного эволюционного древа, цифры, набранные курсивом, — не учтенным. Если число больше 4, это означает, что данный ген претерпел дополнительные дупликации, уже не полногеномные, а тандемные.
Присутствие в геноме мечехвостов дополнительных копий было продемонстрировано не только для Hox-генов, но и для нескольких других групп генов, обычно регулирующих эмбриональное развитие (ParaHox-гены, Six-гены, NK-гены и прочие). Некоторые «избыточные» гены мечехвостов уже превратились в псевдогены, а другие разделили между собой функции, по крайней мере частично (это подтверждено исследованиями областей активности генов у разных видов). Всё это полностью соответствует ожиданиям: именно такие процессы и должны следовать за любой полногеномной дупликацией.
Для очень многих исследованных генов число копий в геноме мечехвостов не ограничивается двумя, а доходит до трех или четырех. Скорее всего, это означает, что событий полногеномной дупликации тут было два — как у позвоночных. Получается, что общий предок современных мечехвостов тоже был октоплоидным.
Когда этот предок жил? Мечехвосты — очень древняя группа животных. Это типичные «живые ископаемые»: хорошо известно, что мечехвосты, жившие 400 миллионов лет назад, отличаются от современных не так уж сильно. Древность этой группы наглядно демонстрируется ее разорванным ареалом: из трех видов мечехвостов, изученных с участием Холланда, один живет вблизи атлантического побережья Северной Америки, а два — на мелководьях морей Юго-Восточной Азии и Китая. На данный момент мечехвостов не назовешь успешной эволюционной ветвью: современных видов в ней всего-то четыре. Судя по молекулярным данным, общий предок современных мечехвостов жил относительно недавно — 135 миллионов лет назад. Но удвоения генома, конечно, могли у них произойти и раньше.
В любом случае, перед нами первый доказанный эпизод крупномасштабной полногеномной дупликации у линяющих животных. Увы, мечехвостам она не принесла такого эволюционного успеха, как позвоночным. Сусуму Оно был прав в том, что генная дупликация является мощным источником эволюционной новизны; но ее одной для прогресса (в каком угодно смысле этого слова) всё же недостаточно. Судя по палеонтологической летописи, даже в эпоху расцвета мечехвостов их было максимум несколько десятков видов — с позвоночными не сравнить. Аналогичный генетический «эксперимент» в двух разных случаях привел к совершенно разным результатам.
Остается неясным, произошло ли удвоение генома только в эволюционной линии мечехвостов или же оно было у общего предка хелицеровых в целом. Кроме мечехвостов, к хелицеровым относится гораздо более многочисленная группа — паукообразные. У некоторых из них, а именно у скорпионов, действительно найдены надежные свидетельства многочисленных генных дупликаций. Но были ли эти дупликации полногеномными — пока сомнительно.
Источник: ЭЛЕМЕНТЫ БОЛЬШОЙ НАУКИ
